Ojos para lo infinitesimal VII: más allá del límite
A finales de 1959, el físico Richard Feynman dio una conferencia a la que tituló Hay mucho espacio al fondo. En ella, entre otras cosas, teorizó acerca de las posibilidades que surgirían si fuésemos capaces de manipular átomos uno a uno, de usar máquinas del tamaño de nanómetros o de cómo se entendería la biología al tener microscopios electrónicos (que usan haces de electrones y no luz en muestras marcadas con átomos pesados) cien veces mejores. Hoy, los microscopios electrónicos tienen resoluciones que les permiten distinguir desde átomos hasta estructuras de unas cuantas de decenas de nanómetros, pero algunos microscopios ópticos ya alcanzan resoluciones cercanas a los veinte nanómetros utilizando luz visible. Sin duda, si el señor Feynman hubiese dado su conferencia hoy habría incluido en su charla a la microscopía óptica que ha superado el límite de difracción.
A finales del siglo XX, una idea que ya llevaba pululando desde que Marvin Minsky conceptualizarse el microscopio confocal tomó dimensiones insospechadas. Esta idea, como ya apuntábamos en Ojos para lo infinitesimal: buscando el límite, era que para incrementar la resolución deberíamos ser capaces de evitar ver la luz que procedía de todo aquello en la muestra que no se quería observar. Porque era el bosque lo que no nos dejaba ver al árbol. Desde entonces, en estos últimos veinte años han ido surgiendo un elenco de tecnologías que encuadran bajo el nombre de microscopía óptica de superresolución. Todas ellas tienen una resolución por encima del límite de difracción calculado por Ernst Abbe en 1873 y ejecutan semejante proeza usando diferentes aproximaciones donde las técnicas de reconstrucción de imagen y la capacidad para modular láseres tienen mucho peso.
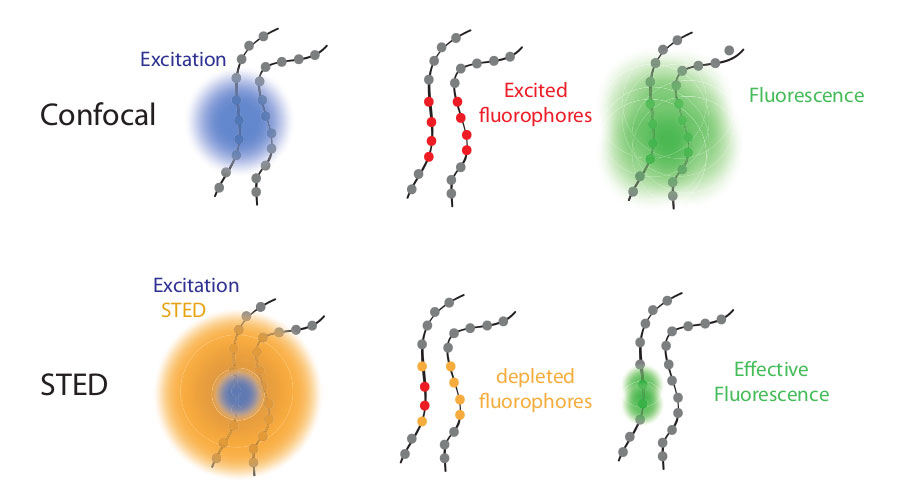
Así por ejemplo una de estas técnicas, denominada STED (por sus siglas en inglés stimulated emission depletion) y desarrollada por Stefan Hell, rompe el límite por medio de un punto focal extraordinariamente pequeño, del tamaño nanómetros. En esa minúscula región de la muestra se enfocan dos láseres. El que ocupa la parte central activa la fluorescencia de las moléculas de fluoróforo en el centro del punto focal mientras que el que se enfoca en la parte externa (formando un donut) evita que las moléculas de esa zona puedan brillar (sí, un láser modulado adecuadamente también puede apagar fluoróforos). De esta manera se escanea la muestra y luego la imagen se reconstruye computacionalmente punto por punto.
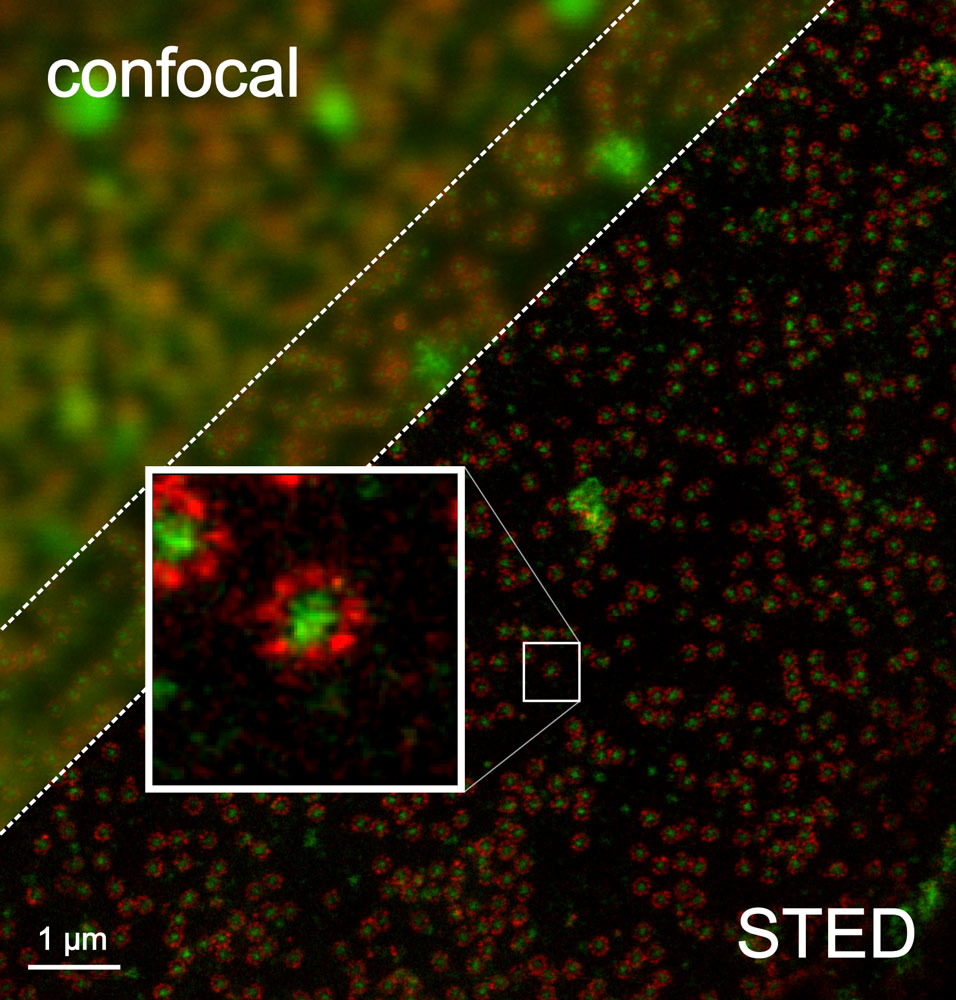
Esquema del funcionamiento de la microscopía STED. Fuente: https://svi.nl/STEDMicroscopyFigura comparación de la superficie de núcleo celular visto desde arriba con microscopio confocal o con un microscopio STED. Marcadas con un fluoróforo rojo proteínas que forman parte de la estructura externa de los poros nucleares (NPC) y con fluoróforo verde se marcan un tipo de proteínas que actúan como puerta en el centro del poro; Fuente: By Tassekaffee - Own work, CC BY-SA 3.0, Wikipedia Commons
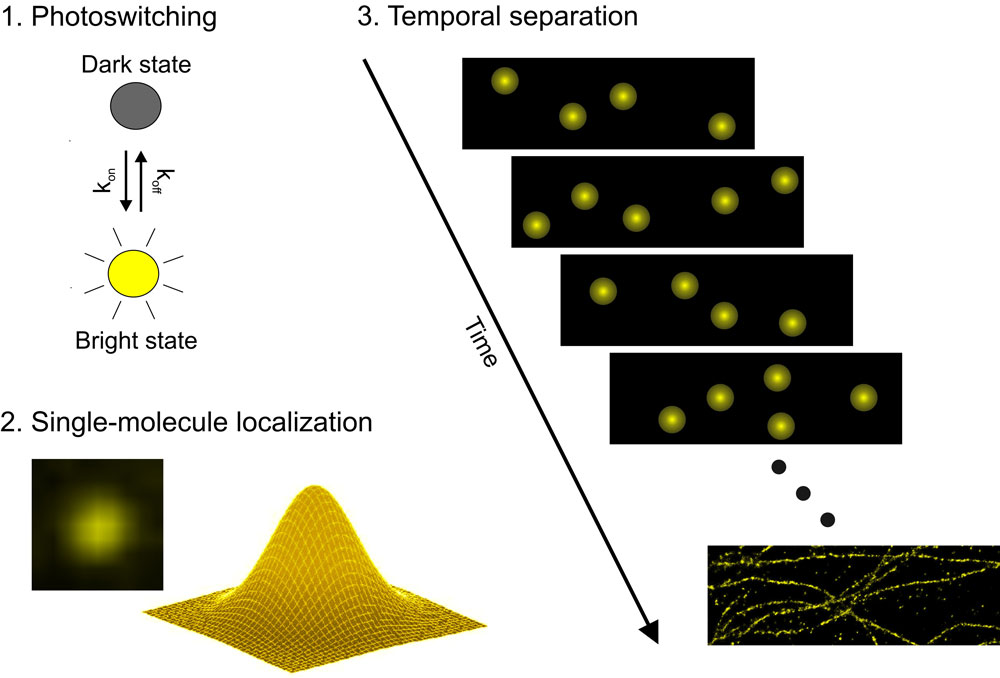
Otros tipos de abordaje para rebasar el límite fueron los usados por los grupos de Xiaowei Zhuang (STORM), o de Samuel Hess (FPALM) o de Eric Betzig y Harald Hess (PALM). Aunque estas tres modalidades son técnicamente diferentes parten de una idea similar: el uso de fluoróforos fotoactivables donde solo unos pocos de ellos son activados y desactivados de manera aleatoria y captados a alta resolución mediante técnicas derivadas de la desarrollada por William E. Moerner a finales de los años 80 para ver moléculas aisladas en disoluciones. Luego, este proceso se repite de manera secuencial hasta que suficientes datos han sido acumulados. Gracias a la aleatoriedad en el proceso de activación de las moléculas debido al uso de haces de luz relativamente débiles, estas señales se encuentran a suficiente distancia como para poder asumir las coordenadas exactas de cada molécula dentro del punto de luz que emite, el cual es decenas de veces más grande que ella. La reconstrucción computacional de la posición exacta de cada molécula en cada secuencia captada y su superposición genera una imagen superresolutiva.
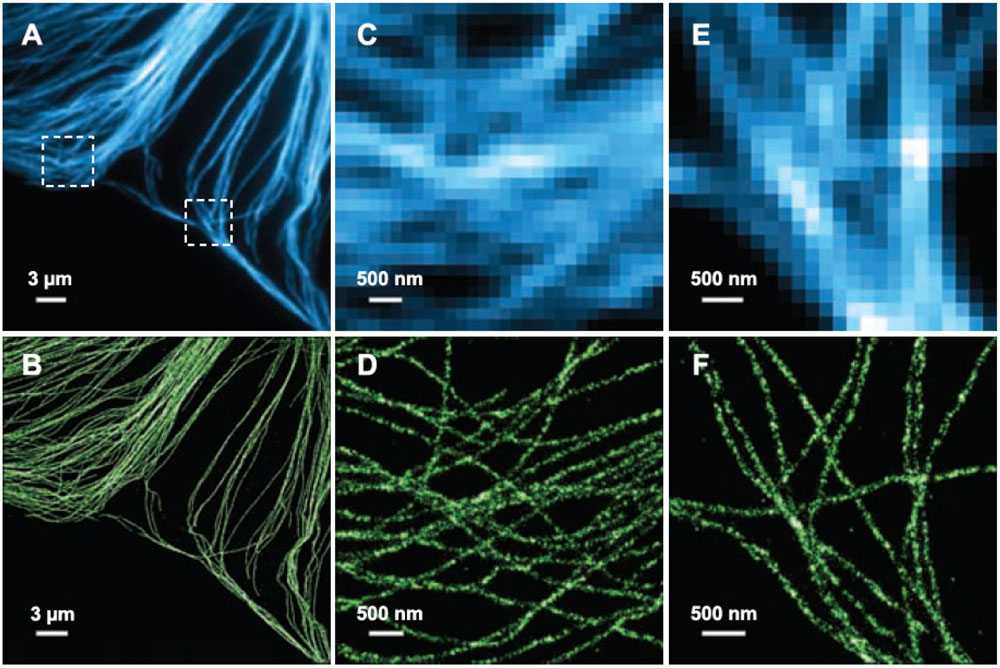
Esquema del funcionamiento de la microscopía STORM. Fuente: https://www.uni-frankfurt.de/49962827/Methods_EquipmentMicrotúbulos en el interior de una célula de mamífero observados con un microscopio convencional (A,C, E) o con un microscopio STORM (B,D,F). Fuente: Bates et al 2007, Science
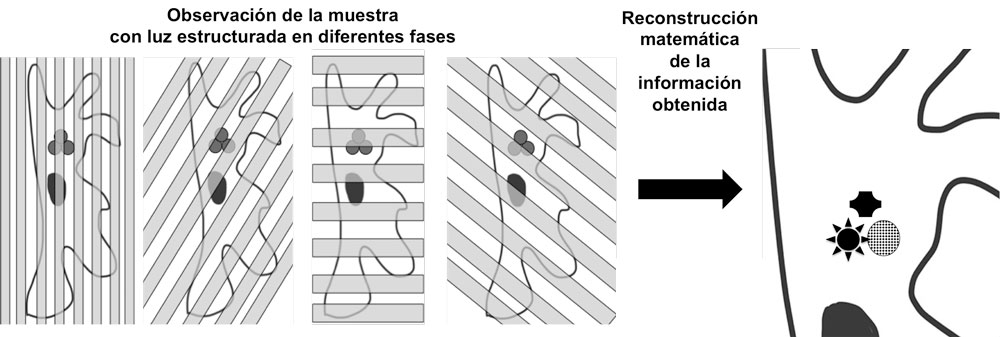
Una aproximación más alejada de las anteriores fue una técnica denominada SIM desarrollada por Mats Gustafsson. En ella se utilizan haces de luz estructurada formando patrones concretos para observar la muestra. La imagen resultante directamente no rompe el límite de resolución, sin embargo, contiene información que puede ser reconstruida computacionalmente mediante funciones matemáticas para generar imágenes que sí se encuentran por encima del límite de resolución en dos dimensiones. Los sucesivos desarrollos han llevado a modalidades de este tipo de microscopía de superresolución que supera el límite de resolución en las tres dimensiones. Y que si bien generalmente no alcanza la resolución de las anteriores, es técnicamente más simple, no usa láseres sino haces de luz no coherente y es más rápida, lo que la hace menos fototóxica y más susceptible de ser aplicada para grabar células vivas completas.
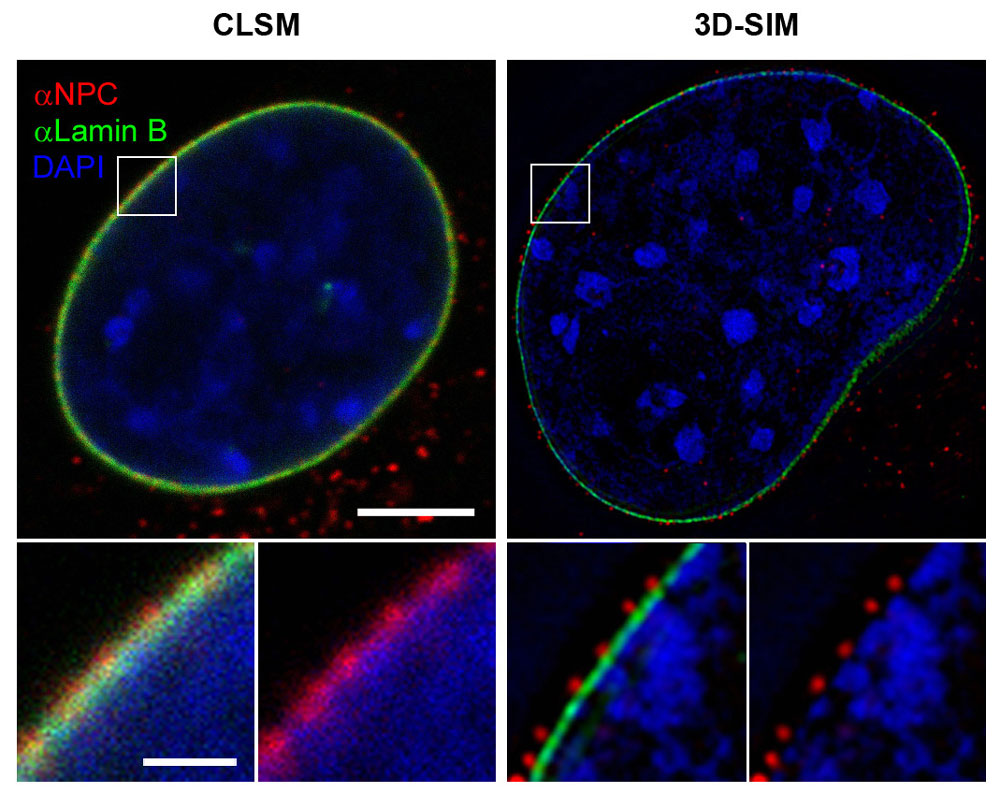
Esquema del funcionamiento de la microscopía SIM. Fuente: Daniel MorenoFigura comparación de un núcleo celular visto en corte transversal con microscopio confocal (CLSM) o con un microscopio SIM con capacidad resolutiva en tres dimensiones (3D-SIM). Marcados con un fluoróforo rojo los poros nucleares (NPC), con fluoróforo verde se marca una proteína de la envoltura nuclear llamada lámina y en azul se marca la cromatina por medio de un compuesto que emite luz azul cuando se une al ADN; Fuente: By Lothar Schermelleh - Lothar Schermelleh, CC BY-SA 3.0, Wikipedia Commons
Actualmente, se cuenta con alrededor de una veintena de variantes y modalidades de microscopía de superresolución, cada una con sus parecidos y diferencias y sus ventajas e inconvenientes. Todas ellas en constante evolución, buscando siempre una mayor resolución espacial y temporal. Yo diría que nuestro objetivo último sería, sabiendo que la existencia de los límites es solo cuestión de tiempo, ser capaces de ver la vida mientras acontece a nivel infinitesimal, proteína por proteína. Por el momento, la resolución espacial máxima alcanzada por alguna de estas técnicas ronda los 20 nanómetros en muestras fijas, lo que está diez veces por debajo de del límite de difracción encontrado por Ernst Abbe y en el rango del tamaño de muchos grupos de proteínas que funcionan formando complejos. Sin embargo, obtener esa misma resolución espacial en grandes volúmenes de muestras vivas (por ejemplo, en una una célula entera) va a ser un poco más complejo. Por ahora solo se han conseguido filmar pequeños volúmenes dentro de la materia viva por unos pocos segundos a elevadísimas resoluciones.
La limitación, por el momento, se encuentra en que las técnicas que alcanzan resoluciones cercanas a los 20 nanómetros necesitan hacer un barrido, escaneo o toma secuencial del volumen que se quiere grabar. Esto tarda un tiempo directamente proporcional al volumen en cuestión. Si el volumen es pequeño el tiempo para cada fotograma será corto y las moléculas de la materia viva no se moverán demasiado, generando fotogramas de alta resolución. Por el contrario, si el volumen es grande el tiempo para cada fotograma será largo y las moléculas de la materia viva tendrán tiempo para desplazarse mucho, generando fotogramas borrosos poco resolutivos.
Sin embargo, existen también otros tipos de microscopía desarrollados recientemente denominados de «hoja de luz» que usan haces de luz estructurada (diferentes a los comentados anteriormente). Estos son capaces de grabar grandes volúmenes de espacio (en los que caben desde células solitarias hasta minúsculos embriones de insectos o peces) en milésimas de segundo con una resolución espacial cercana al límite de difracción, aunque sin superarlo.
En el siguiente enlace se puede ver un video de alta resolución de los cromosomas marcados con proteína roja fluorescente (RFP) y el huso miótico marcado con GFP de una célula en diferentes fases de la mitosis. En este otro enlace tenemos un video de alta resolución de una célula (marcada de naranja) durante su migración vista desde diferentes ángulos.
Así pues, como empujados por esa virtud que ha seguido con paciencia a la especie humana durante milenios, límite tras límite y después de siglos de gigantes apoyados a hombros de gigantes, conseguimos por fin ver con nuestros ojos en las entrañas de la vida a escalas que se creían otrora imposibles. Va a ser fascinante ver dónde nos llevan nuestros nuevos y mejorados ojos para lo infinitesimal.
Referencias:
—Light Microscopy: 1959, Invention of the microscope. Nature Milestones (2009) —Nobel Prize for Stanford chemist W.E. Moerner, who brings very small things into focus. Stanford News (2014). —Bates, M., Huang, B., Dempsey, G. T. & Zhuang, X. Multicolor Super-Resolution Imaging with Photo-Switchable Fluorescent Probes. Science (80-. ). 317, 1749–1753 (2007). —Chen, B.-C. et al. Lattice light-sheet microscopy: Imaging molecules to embryos at high spatiotemporal resolution. Science (80-. ). (2014). doi:10.1126/science.1257998 —Shao, L., Kner, P., Rego, E. H. & Gustafsson, M. G. L. Super-resolution 3D microscopy of live whole cells using structured illumination. Nat. Methods 8, 1044–1046 (2011). —Saxena et al. Structured illumination microscopy. Advances in Optics and Photonics 7, 241–275 (2015).



Deja tu comentario!